Resúmenes de articulos
Se muestran seis ejemplos desde el punto de vista calorimétrico y termodinámico de sistemas vivos. Anteriormente filósofos se dieron cuenta que los animales no podían sobrevivir en una atmósfera donde no existiera una flama.Los organismos vivos son sistemas abiertos en constante intercambio de materia, energía entropía e información con su ambiente que mueren cuando son aislados en un sistema cerrado aproximado a un equilibrio térmico.
Los organismos degradan glucosa anaeróbicamente para obtener moléculas de ATP.Queda claro que la producción bioenergética de calor permanece estable cuando la radiación y la evaporación disminuyen radicalmente.Un ejemplo de la termodinámica en seres vivos son los enjambres de abejas, una abeja sola a 20°C tiene una masa térmica de 208mWg-1 pero este rango disminuye hiperbólicamente cuando aumenta el número de abejas, Los músculos de vuelo tienen el mayor cambio energético, existen abejas con un sistema de defensa muy peculiar, cuando una avispase acerca al panal recibe advertencias inútiles de las abejas, es entonces cuando un grupo de 400 abejas se lanza sobre la avispa formando una esfera alrededor de ella, generando en el centro temperaturas de 46°C, debido a la tasa metabólica de las abejas, quemando prácticamente a la avispa. La energía consumida por las abejas es de 9kj, que equivale a 0.6g de miel, lo que hace a este sistema muy efectivo.El uso de la relación entre la respiración y la producción de calor puede ser correlacionado con la salud y con distorsiones de esta relación puede ser relacionadas con enfermedades o debilidad, siendo de gran utilidad en el diagnostico o supervisión.
El balance energético puede ser encontrado en el desarrollo de algunos escarabajos e insectos donde el estado larval necesita recolectar la mayor cantidad de energía para el estado de pupa.
Propone que la primer transducción de energía se dio a partir de la oxidación de sulfuro de hidrogeno, y sulfuro férrico a pirita y dos protones, resultando en un gradiente de protones a través de la membrana, que permitiría una fuerza protón motriz.La vida tiene dos características clave, la propagación de información y la transducción de energía.
Los reactivos originales presentes en dentro y fuera de la célula primitiva reaccionaron con un complejo metálico asociado a la membrana de la célula, similar a lo que sucede con las células de hoy en día, los electrones son transmitidos por compuestos organometalicos liposolubles.
Posiblemente estén involucrados metales en transición y compuestos iónicos de sulfuro, similares a las ferrodoxinas y CO reductasas, así como ATPasas generando compuestos fosforilados con la fuerza protón motriz.
Como los compuestos reductores abundarían y debido a las condiciones ambientales, la energía libre estándar seria negativa.
Un árbol filogenético confirma las divisiones monofileticas entre eucariontes, procariontes y arqueobacterias establecidas por RNAs.
Una estructura membranal cerrado es importante, así como la reacción de formación de pirita o fuentes de protones y e- se realizarían fuera de la célula.
Cada organismo vivo podría contener remanentes de los procesos energéticos más primitivos, pues todos generan fuerza protón motriz a través de una cadena de electrones.
Debido al enfriamiento de la tierra, donde el agua líquida seria abundante, la reducción de CO y CO2 con reductores más fuertes que H2, sería posible en un ambiente liquido.
La pregunta es ¿el estado vivo surge necesariamente de estructuras celulares preexistentes que requirió para capturar energía para procesos de crecimiento? . Se han encontrado meteoritos de la corteza marciana con inclusiones de carbonato, siendo interpretado como un origen bioenergético.
Probablemente la materia orgánica haya sido traída a la superficie terrestre desde un meteorito. La materia orgánica ha ido desapareciendo a diferentes ritmos, por una serie de reacciones como la hidrólisis y pirolisis.
Por el contrario parece ser que los sistemas vivos se desarrollaron en soluciones diluidas de algunos sustratos como aminoácidos, carbohidratos, y bases puricas. Considerando que la forma más simple de captura de energía fue por sistemas moleculares de distintas formas, como la condensación por deshidratación. Una segunda forma de captura es por reacciones de oxido reducción, teniendo a gradientes iónicos como fuentes potenciales de energía.
Los hidrocarburos y sus derivados estuvieron presentes en la reserva orgánica de la tierra primitiva, permitiendo la formación de membranas por organismos primitivos.
Erwin Schrödinger en una conferencia pregunta ¿por qué los átomos son tan pequeños?, Los procesos físicos y químicos básicos para el funcionamiento de un organismo requieren la participación de grandes cantidades de átomos para realzarse de modo seguro y previsible, por lo que una estructura con pocos átomos no podría permitir procesos biológicos elementales.
Un organismo vivo se mantiene ordenado a pesar de la agitación térmica y otros comportamientos que tienden a desordenar cualquier estructura constituida por muchas partículas, si es así, ¿por qué se puede crear y mantener el orden de un ser vivo?
Erwin Schrödinger responde de dos maneras, orden a partir del desorden y orden a partir del orden, la primera explica que un organismo se ordena por una pérdida del orden del ambiente, la según que el orden se transmite genéticamente; el ADN almacena información y se mantiene estable.
Max Delbrück, describe que moléculas grandes formadas por uniones covalentes tienen suficiente estabilidad como para almacenar información genética.
Finalmente opina que la vida es un proceso diferente único que será explicado por las leyes físicas.
La biología será útil a la física permitiendo descubrir nuevas leyes, y también la física a la biología ofreciendo una explicación unificada de la vida
Un radical libre es cualquier molécula que posea un solo electrón desapareado.
El proceso de tomar electrones de otros átomos se llama oxidación, la sustancia que toma los electrones se reduce.
El resultado de la oxidación d de un electrón del oxigeno es el radical superóxido O_2^(∙-) , si dos electrones se transfieren el producto es peróxido de hidrogeno, H2O2, que no es un radical, el oxido ferroso es capaz de transferir un tercer electrón al H2O2 produciendo la lisis del enlace O-O, una parte se reduce a agua y la otra es el radical hidroxilo, HO∙ un oxidante muy poderoso, que incluso puede ser agente bactericida, la única enzima que produce este radical intencionalmente es la nicotinamina ADP oxidasa que se encuentra en los neutrofilos.
El radical hidroxilo puede desencadenar la peroxidación e lípidos e incluso atacar y romper al ADN.
Se demuestra que el hierro acelera el estrés oxidativo, indicando que los hombres acumulan hierro casi linealmente desde la pubertad, al contrario de las mujeres que están protegidas hasta la menopausia, sin embargo no existe un mecanismo para eliminar el exceso de hierro.
Las vitaminas antioxidantes, E y C, ayudan a terminar la peroxidación en cadena de lípidos.
El olfato junto a otros sentidos vela por nosotros dando a nuestro cerebro la información necesaria para movernos en el mundo que nos rodea. El olfato es el sentido más primitivo y el primero en aparecer en la escala evolutiva, tras aparecer en los peces el olfato se separa anatómicamente del gusto en los anfibios, hace 400 millones de años.
Las plantas utilizan el olor para atraer insectos, los insectos como las hormigas reconocen y se orientan por el olor y los mosquitos seleccionan a sus presas también por el olor.los machos de algunas mariposas perciben el olor de las hembras a kilómetros, y otros animales utilizan el olfato para reconocer lugares, localizar presas o incluso a sus depredadores.
Las principales causas de pérdida del olfato son el resfriado, inflamación de la mucosa, traumatismos craneofaciales, tabaquismo y enfermedades neurodegenerativas.
La historia del nobel
Linda Buck y Richard Axel observaron que el ARN obtenido del epitelio olfativo de rata contenía una gran familia de genes entre 70 y 200, después secuenciaron las proteínas de estos genes , obteniendo una serie de proteínas con 7 dominios transmembranales. Estas proteínas tenían una estructura 7-transmebrana y la activación de estas producía una cascada de eventos desde la activación de proteínas g generando un aumento en AMPc y así activar a neuronas olfativas. Recibiendo el premio nobel de medicina o fisiología en 2004.
Olfato y gusto son parte de sistema sensorial químico, permiten la interpretación de olores y sabores, determinan la supervivencia diaria de muchas especies.
Desarrollar un olfato artificial es fundamental, para que permita detectar y cuantificar olores de forma analítica y con precisión al igual que un sentido del gusto.
Olores son mezcla de multitud de moléculas, la relación es entre estructura molecular y olor
Los más olorosos son sustancias con estructuras en anillo con electrones no localizados, estructuras “aromáticas”.
Tiene una estimulación exclusiva a través de moléculas olorosas, que llegan a un epitelio olfativo que tiene una capa mucosa con neuronas olfativas ciliadas.
En los cilios comienza la transducción, moléculas olorosas se “acoplan” a proteínas receptoras tipo G, que activa a la adenilato ciclasa catalizando la formación de AMPc, logrando una apertura en canales de Na+, despolarizando la membrana, llevándose una amplificación de la señal.
A grandes rasgos podemos distinguir entre dos tipos de información, las que van a zonas superiores de procesamiento olfativo y aquellas que van a estructuras límbicas, las primeras relacionadas con el reconocimiento consciente de olores y la segunda con las respuestas subconscientes a olores y la relación con emociones.
El epitelio olfativo no tiene zonas definidas para un cierto tipo de molécula olorosa.
Si un glomérulo recibe la información de células que poseen un cierto receptor entonces el glomérulo situado en la misma posición en el bulbo del otro lado también recibirá la misma información. Localizando en los distintos animales la posición especifica conservada.
El gusto.
La lengua es el principal órgano gustativo del cuerpo humano, recu8bierto por papilas de cuatro tipos: fungiformes, circunvaladas, foliadas, y filiformes.
Las patologías del gusto y el olfato se clasifican en tres:
Anosmia, se pierde la capacidad de detectar sabores u olores, hiposmiala capacidad de detectar olores o sabores de un tipo se ve afectada, y trastornos de la percepción donde se perciben olores o sabores que no existen en realidad.
Sin embargo no has trabajos que permitan un tratamiento a estas patologías.
El olfato artificial.
Definición:
Una nariz electrónica es un instrumento que consiste en un conjunto de sensores electroquímicos con una especificidad parcial y un sistema adecuado de reconocimiento de patrones, capaz de reconocer olores simples o complejos.
Nada antes de 1920. En 1920 se postularon primeras teorías y a mediados de los 60 aparecen sensores de conductividad.
Una nariz electrónica es un sistema que consiste en: un suministro de la muestra o material oloroso, una cámara con un arreglo de sensores, un sistema de procesamiento de las señales, un sistema conversor A/D, un sistema de análisis y un sistema de visualización de resultados.
Las aplicaciones para la nariz electrónica son varias, automoción, salud, control de entorno, sector farmacéutico, perfumería, seguridad y aplicaciones similares.
Gusto artificial
La lengua artificial es un instrumento analítico que reproduce de forma artificial la sensación del sabor. Constan de un muestreador, un conjunto de sensores químicos un instrumento para adquirir la señal, y el software para procesar la información. Funciona similar a la nariz electrónica, pero le medio de detección es liquido.
Cuestionario
Es la que estudia los procesos en los que la energía se transfiere como calor y trabajo.
El calor es muy parecido al trabajo, para distiguirlos el calor se define como una transferencia de energía provocada por una diferencia de temperatura, mientras que el trabajo es una transferencia de energía que no se debe a una diferencia de temperatura.
En la termodinámica, con frecuencia se hace referencia a sistemas particulares. Un sistema es cualquier objeto o conjunto de objetos que se desean considerar, al resto del universo se le considera como el ambiente o los alrededores.
2. ¿A qué se refiere la constancia de equilibrio?
Cuando en un sistema químico la cantidad de moléculas de producto como las de reactivo son constantes, a demás de que la velocidad de ambas (productos y reactivos) se iguala.
3. ¿Qué es el ATP?
Por su siglas en inglés: adenosín triphosfato es un nucleótido fundamental en la obtención de energía celular. Está formado por una base nitrogenada (adenina) unida al carbono 1 de un azúcar de tipo pentosa, la ribosa, que en su carbono 5 tiene enlazados tres grupos fosfato. Se encuentra incorporada en los ácidos nucleicos.
Se produce durante la fotosíntesis y la respiración celular, y es consumido por muchas enzimas en la catálisis de numerosos procesos químicos.
4. Al hidrolizarse el ATP, cuando se rompe un enlace, ¿cuánta energía libera?
ΔG = -7,7 kcal/mol
5. ¿Qué es el potencial redox?
También es llamado potencial de reducción es un concepto electroquímico que se puede determinar midiendo la actividad de los electrones.
6. ¿Qué es un radical libre?
Son átomos o moléculas extremadamente reactivas, debido a que en el orbital más externo de su estructura tienen uno o más electrones sin aparear. Esta inestabilidad les confiere una necesidad física por la captura de un electrón de cualquier otra molécula de su entorno, ocasionando que la estructura afectada quede inestable. De esta forma pueden establecer reacciones en cadena por medio de varios transportadores que se oxidan y se reducen secuencialmente.
7. ¿De qué habla la teoría quimiosmotica?
Propuesta por Mitchell, explica cómo la energía derivada del transporte de electrones por la cadena de transporte de electrones se utiliza para producir ATP a partir de ADP y Pinorganico. La bomba de protones: el transporte de electrones está acoplado al transporte de H+ a través de la membrana interna mitocondrial desde el espacio intermembranal. Este proceso crea simultáneamente a través de la membrana interna mitocondrial un gradiente eléctrico (con más cargas positivas en el exterior de la membrana que en la matriz mitocondrial) y un gradiente de pH (el exterior de la membrana está a un pH más ácido que el interior). La energía generada por este gradiente es suficiente para realizar la síntesis de ATP.
8. ¿Cuáles son los inhibidores de la respiración?
Este tipo de inhibidores reciben este nombre porque su principal función es el inhibir el transporte de electrones en la cadena de la respiración. Los inhibidores del transporte de electrones más comúnmente usados pueden reunirse en tres grupos principales según el sitio de la cadena respiratoria donde actúan:
1.Sobre la NADH-deshidrogenasa, bloqueando la transferencia de electrones entre la flavina y la ubiquinona. (Inhibidores del sitio I)
• Barbitúricos, como el amobarbital
• Piericidina A (antibiótico)
• Rotenona (insecticida)
2.Actúa bloqueando la transferencia de electrones entre el citocromo b y el citocromo c1. (inhibidores de sitio II)
3.Actúan sobre el Hemo a3 de la citocromooxidasa impidiendo su interacción con el oxígeno (inhibidores de sitio III)
• Cianuro
• Monóxido de carbono
• H2S
9. Menciona algunas enfermedades genéticas causadas por DNAmt
Desórdenes motores, accidentes cerebrovasculares, convulsiones, demencia, intolerancia al ejercicio, oftalmoplejia, retinopatía pigmentaria, atrofia óptica, ceguera, sordera, cardiomiopatía, disfunciones hepáticas y pancreáticas, diabetes, defectos de crecimiento, anemia sideroblástica, pseudo obstrucción intestinal, nefropatías, acidosis metabólica y otras más secundarias.
10. ¿Qué es la fotoquímica?
La fotoquímica es el estudio de las transformaciones químicas provocadas o catalizadas por la emisión o absorción de luz visible o radiación ultravioleta. Una molécula en su estado fundamental (no excitada) puede absorber un quantum de energía lumínica, esto produce una transición electrónica y la molécula pasa a un estado de mayor energía o estado excitado. Una molécula excitada es más reactiva que una molécula en su estado fundamental.
11. ¿Cuáles son los mecanismos que participan en el transporte de las moléculas a través de las membranas?
El transporte que requiere la acción de proteínas transportadoras específicas en la membrana denominado transporte con intervención de transportador, este a su vez se puede dividir en difusión facilitada y transporte activo
El transporte a través de la membrana sin intervención de transportadores, este implica a la difusión simple de iones.
12. ¿Cuál es el espectro de luz que los humanos pueden ver?
El espectro de luz visible que va desde 400 hasta los 750 nm osea desde el violeta hasta el rojo
13. ¿Qué son los conos y bastones?
Son dos tipos de neuronas fotorreceptoras y ambas células contienen moléculas de pigmento que se disocian como respuesta de luz y esta reacción fotoquímica da lugar finalmente a la producción de potenciales de acción en el nervio óptico. Los bastones proporcionan la visión en blanco y negro en condiciones de intensidad baja de luz, mientras que los conos proporcionan una visión fina de color cuando las intensidades luminosas son mayores.
14. ¿Qué es el sabor?
El sabor lo provoca receptores constituidos por papilas gustativas con forma cilíndrica, localizadas principalmente en la superficie dorsal de la lengua, cada una de estas papilas gustativas están constituidas por 50 – 100 células epiteliales especializadas que presentan microvellosidades largas que se extienden a través de un poro situado en la propia papila hasta que alcanzan el medio externo donde se bañan en saliva. Aunque estas células epiteliales sensitivas no son neuronas, se comportan como tal; se despolarizan cuando se estimulan adecuadamente, producen potenciales de acción y liberan neurotransmisores que estimulan a las neuronas sensitivas relacionadas con las papilas gustativas.
15. ¿Cuáles son las bases moleculares de la olfacción?
Las moléculas que inducen olor se unen a receptores y actúan a través de proteínas G para incrementar el AMPc en el interior de la célula. A su vez este incremento da lugar a la apertura de canales de membrana y causa la despolarización del potencial generador que, por su parte, estimula la producción de potencial de acción. Cada proteína receptora se puede asociar a un máximo de 50 proteínas G. la disociación de estas proteínas G da lugar a la liberación de numerosas subunidades de proteína G, lo que amplifica el efecto en muchas veces. Esta amplificación puede explicar la gran sensibilidad del sentido del olfato: la nariz del ser humano puede detectar la millonésima parte de un gramo de perfume en el aire, aun así nuestro sentido del olfato es mucho más grosero que el de otros mamíferos.
Olfato
Cada neurona bipolar tiene una dendrita que se proyecta hasta la cavidad nasal, en donde finaliza formando una protuberancia que contiene un cilio. La neurona sensitiva bipolar también presenta un único axón amielínico que se proyecta a través de una serie de orificios que existen en la placa cribiforme del etmoides, hasta el bulbo olfativo del cerebro, en donde estable sinapsis con neuronas del segundo nivel. Por tanto, a diferencia de otras modalidades sensitivas que alcanzan el cerebro a partir del tálamo, el sentido del olfato se transmite directamente hasta la corteza cerebral. El procesamiento de la información olfativa comienza en el bulbo olfativo en el que las neuronas sensitivas bipolares hacen sinapsis con las neuronas localizadas en tramas esféricas denominadas glomérulos. Existen pruebas que indican que cada uno de estos glomérulos recibe aferencias de un solo tipo de receptor olfativo. El olor de una flor, que libera muchas moléculas olorosas diferentes, se puede identificar por el patrón de excitación a que da lugar en los glomérulos del bulbo olfativo. La identificación de un olor mejora por la inhibición lateral en el bulbo olfativo, que parece implicar la existencia de sinapsis dendrita-dendrita entre las neuronas de los glomérulos adyacentes.

Las neuronas del bulbo olfativo se proyectan hasta la corteza olfativa en la parte medial de los lóbulos temporales, y también hasta otras estructuras relacionadas como el hipocampo y el núcleo amigdalino. Estas estructuras forman parte del sistema límbico y que desempeñan un papel importante en las emociones y la memoria. En concreto, el núcleo amigdalino humano ha sido implicado en las respuestas emocionales frente a la estimulación olfativa y quizá esta sea la razón por la que la percepción de un olor concreto puede evocar de manera tan intensa recuerdos cargados de emociones.
Las bases moleculares de la olfacción son complejas. Al menos en algunos casos, las moléculas que inducen olor se unen a receptores y actúan a través de proteínas G para incrementar el AMPc en el interior de la célula. A su vez este incremento da lugar a la apertura de canales de membrana y causa la despolarización del potencial generador que, por su parte, estimula la producción de potencial de acción. Cada proteína receptora se puede asociar a un máximo de 50 proteínas G. la disociación de estas proteínas G da lugar a la liberación de numerosas subunidades de proteína G, lo que amplifica el efecto en muchas veces. Esta amplificación puede explicar la gran sensibilidad del sentido del olfato: la nariz del ser humano puede detectar la millonésima parte de un gramo de perfume en el aire, aun así nuestro sentido del olfato es mucho más grosero que el de otros mamíferos.
Se ha descubierto una familia de genes que codifican las proteínas del receptor olfativo. Es una gran familia que puede incluir hasta mil genes. Este gran número puede reflejar la importancia del sentido del olfato en los mamíferos en general. Sin embargo, la existencia de mil genes diferentes que codifican un millar de proteínas receptoras también diferentes no puede explicar el hecho de que el ser humano pueda diferenciar más de 10 000 olores diferentes. Claramente, el cerebro debe realizar la integración de las señales procedentes de varias neuronas sensitivas que presentan proteínas receptoras olfativas diferentes, interpretando después el patrón como una “huella dactilar” característica de cada olor concreto.
Texto e imágenes: Fox, Stuart Ira. Fisiología humana séptima edición McGraw-Hill interamericana 2003, pp. 254-255
Gusto
El sabor, correspondiente al sentido del gusto, lo provoca receptores constituidos por papilas gustativas con forma cilíndrica, localizadas principalmente en la superficie dorsal de la lengua, cada una de estas papilas gustativas están constituidas por 50 – 100 células epiteliales especializadas que presentan microvellosidades largas que se extienden a través de un poro situado en la propia papila hasta que alcanzan el medio externo donde se bañan en saliva. Aunque estas células epiteliales sensitivas no son neuronas, se comportan como tal; se despolarizan cuando se estimulan adecuadamente, producen potenciales de acción y liberan neurotransmisores que estimulan a las neuronas sensitivas relacionadas con las papilas gustativas.
Existen 4 modalidades principales de sabor o gusto, cada una de las cuales se percibe con mayor agudeza en una región correcta de la lengua. Estos sabores son dulce (en la punta de la lengua), ácido (a los lados de la lengua), amargo (parte dorsal de la lengua) y salado (en la mayor parte de la lengua pero sobre todo en los lados). Todos los sabores diferentes que pueden percibir constituyen combinaciones de estos 4, junto con el efecto aportado por el sentido del olfato. Existe evidencia de que el ser humano presenta un quinto tipo de receptor específico del gusto que ha denominado umami (por glutamato monosódico). También se ha sugerido que para el ser humano puede presentar una modalidad distinta del gusto para el agua.

El sabor salado del alimento se debe a la presencia de iones sodio Na+ o de alguno de otros cationes que activan de manera específica las células receptoras del sabor salado. El Na+ se introduce en las células receptoras sensitivas a través de canales localizados en las membranas apicales. Así tiene lugar la despolarización de las células, con la liberación de su neurotransmisor. Sin embargo, el anión asociado al Na+ modifica de manera sorprendente la intensidad del sabor salado percibido: el NaCl tiene un sabor mucho más salado que otras sales de sodio como el acetato de sodio. Existen pruebas que indican que los aniones pueden pasar a través de las uniones estrechas entre las células receptoras, y que el anión Cl¯ pasa a través de esta barrera con mayor facilidad que los otros aniones. Posiblemente esta diferencia está relacionada con la capacidad del Cl¯ para dar un sabor más salado al Na+, en comparación con los otros aniones.
El sabor ácido, al igual que el sabor salado, se debe al movimiento de iones a través de canales de membrana. Sin embargo el sabor ácido, se debe a la presencia de iones hidrógeno H+, por lo tanto todos los ácidos tienen un sabor ácido.
Al contrario de lo que ocurre con los sabores salado y ácido, los sabores dulce y amargo se deben a la interacción de las moléculas del gusto con proteínas receptoras de membranas específicas.
La mayoría de las moléculas orgánicas, sobre todos los azúcares, tienen un sabor dulce de grados diferentes. El sabor amargo lo provoca la quitina y otras moléculas aparentemente no relacionadas. El sabor amargo es la sensación gustativa más aguda y se suele asociar a moléculas tóxicas (aunque no todos los tóxicos tienen un sabor amargo). Las sensaciones dulces y amargas se producen a través de los receptores acoplados a proteínas G (asociación de tres subunidades de proteínas de la membrana denominadas alfa, beta y gamma, regulada por nucleótidos de guanosina GDP y GTP. Las subunidades de la proteína G se disocian en respuesta a señales de la membrana y, a su vez, activan a otras proteínas celulares). El tipo concreto de proteína G implicado en el sabor ha sido identificado recientemente y denominado gustducina. Este término se ha utilizado para ser hincapié en la similitud con un grupo relacionado de proteínas G de un tipo denominado transducina asociado a los fotorreceptores del ojo. La disociación de la subunidad de proteína G gustducina activa sistemas de segundo dando lugar a la despolarización de la célula receptora, que a su vez, la célula receptora estimulada activa una neurona sensitiva asociada que transmite impulsos hasta el cerebro, en donde se interpreta como la correspondiente percepción gustativa.
Aunque todos los receptores de los sabores dulce y amargo actúan a través de proteínas G, los sistemas de segundo mensajero activados por estas proteínas G dependen de la molécula que induce la percepción sensitiva. Por ejemplo, en el caso del sabor dulce de los azúcares, las proteínas G activan al adenilato ciclasa produciendo AMP cíclico. A su vez, el AMPc produce una despolarización mediante el cierre de canales para el K+ que estaban abiertos previamente. Por otra parte el sabor dulce de los aminoácidos fenilalanina y triptófano, así como el de los edulcorantes artificiales sacarina y ciclamato, pueden activar otros sistemas distintos de segundo mensajero. En este proceso se incluye la activación de la enzima de membrana que da lugar a la aparición de los segundos mensajeros inositol trifosfato IP3 y diacilglicerol DAG.
Texto e imágenes: Fox, Stuart Ira. Fisiología humana séptima edición McGraw-Hill interamericana 2003, pp. 252-254
Visión

Uno ve un objeto en una de dos formas:
1. Cuando el objeto es una fuente de luz, una ve la luz emitida directamente como una llama.
2. Uno ve un objeto por luz que se refleja en el.
La luz visible es aquella a la que los ojos son sensibles y consta de frecuencias que van desde 4x1014 Hz hasta 7.5x1014 Hz, que corresponden a las longitudes de onda en el aire de aproximadamente 400 a 750 nm. A esto se le reconoce como espectro visible y dentro de él se encuentran los diferentes colores desde el violeta hasta el rojo como se observa en la fotografía

El ojo es el órgano presente en algunos animales que permite la detección de cambios de luz y la capacidad de transformar éstos en impulsos eléctricos. Existen dos tipos de neuronas fotorreceptoras que son los conos y bastones. Ambas células contienen moléculas de pigmento que se disocian como respuesta de luz y esta reacción fotoquímica da lugar finalmente a la producción de potenciales de acción en el nervio óptico. Los bastones proporcionan la visión en blanco y negro en condiciones de intensidad baja de luz, mientras que los conos proporcionan una visión fina de color cuando las intensidades luminosas son mayores.
La fotoquímica de la visión comienza los fotorreceptores (conos y bastones) que se activan cuando la luz da lugar a una modificación química de las moléculas del pigmento contenido en el interior de las laminillas membranosas de los segmentos externos de las células receptoras.
Los bastones contienen un pigmento púrpura denominado rodopsina (una combinación de rojo y azul) debido a que transmite la luz en regiones rojas y azul del espectro, al tiempo en el que absorbe la energía luminosa en la región verde. En condiciones de obscuridad, que es cuando se utilizan los bastones para la visión, se ven con más facilidad los objetos verdes que los rojos. La razón es que la luz roja no se absorbe por la rodopsina y solo la luz absorbida puede dar lugar a la reacción fotoquímica que produce la visión. En respuesta a la luz absorbida, la rodospina se disocia en 2 componentes: el pigmento retinaldehído, que procede de la vitamina A, y una vitamina denominada opsina. Esta reacción se denomina reacción de decoloración.
Los conos tienen una sensibilidad frente a la luz menor que los bastones pero los conos proporcionan la visión del color y facilitan una mayor agudeza visual. El ser humano y otros primates presentan una visión tricromática del color, esto quiere decir que nuestra percepción de una gran cantidad de colores se debe a la estimulación de solo tres tipos de conos (azules, verdes y rojos)
La percepción es un proceso nervioso superior que permite al organismo, a través de los sentidos, recibir, elaborar e interpretar la información proveniente de su entorno.
Texto e imágenes: Fox, Stuart Ira. Fisiología humana séptima edición McGraw-Hill interamericana 2003, pp. 266-279
http://farm1.static.flickr.com/196/518431221_19dbbd2010.jpg
Transporte a través de membranas
Por lo tanto se dice que la membrana plasmática es de permeabilidad selectiva. En general, la membrana plasmática no es permeable a las proteínas, los ácido nucleicos y otras moléculas necesarias para la estructura y función de la célula. Sin embargo, es permeable a muchas otras moléculas, permitiendo un tráfico de nutrientes y desechos en dos direcciones necesario para sostener el metabolismo. La membrana plasmática es también selectivamente permeable a determinados iones; esto permite corrientes electroquímicas a través de la membrana, que se utilizan para producir impulsos en las células nerviosas y musculares.
Los mecanismos que participan en el trasporte de moléculas e iones a través de la membrana celular se pueden dividir en dos tipos:
1. El transporte que requiere la acción de proteínas transportadoras específicas en la membrana denominado transporte con intervención de transportador, este a su vez se puede dividir en difusión facilitada y transporte activo
2. El transporte a través de la membrana sin intervención de transportadores, este implica a la difusión simple de iones.
Los procesos de transporte se pueden clasificar por sus requerimientos energéticos. El transporte pasivo es el de movimiento neto de moléculas e iones a través de una membrana desde una concentración más alta a una inferior (a favor de un gradiente de concentración), no requiere energía metabólica.
El transporte pasivo comprende la difusión simple, la osmosis y la difusión facilitada. El transporte activo es un movimiento neto a través de una membrana que se produce contra un gradiente de concentración. El transporte activo requiere el gasto de energía metabólica ATP e implica proteínas trasportadoras específicas.
Los iones inorgánicos como Na+ y K+ son capaces de atravesar la membrana a través de poros existentes en el seno de las proteínas integrales que abarcan la totalidad del espesor de las capas dobles de fosfolípidos.
Con el fin de mantener el metabolismo, las células deben captar glucosa, aminoácidos y otras moléculas orgánicas del medio extracelular. Sin embargo, estas moléculas son demasiado grandes y polares para atravesar la barrera lipídica de la membrana plasmática por un proceso de difusión simple. El transporte de estas moléculas se produce por proteínas trasportadoras de membrana.
Difusión facilitada
Impulsada por energía térmica de las moléculas que difunden e implica un transporte neto desde el lado mayor concentración al de menor concentración.

Transporte Activo
Movimiento de moléculas e iones en contra de sus gradientes de concentración, es decir, desde concentraciones más bajas hasta más elevadas. Este transporte requiere el consumo de energía celular obtenida a partir de ATP
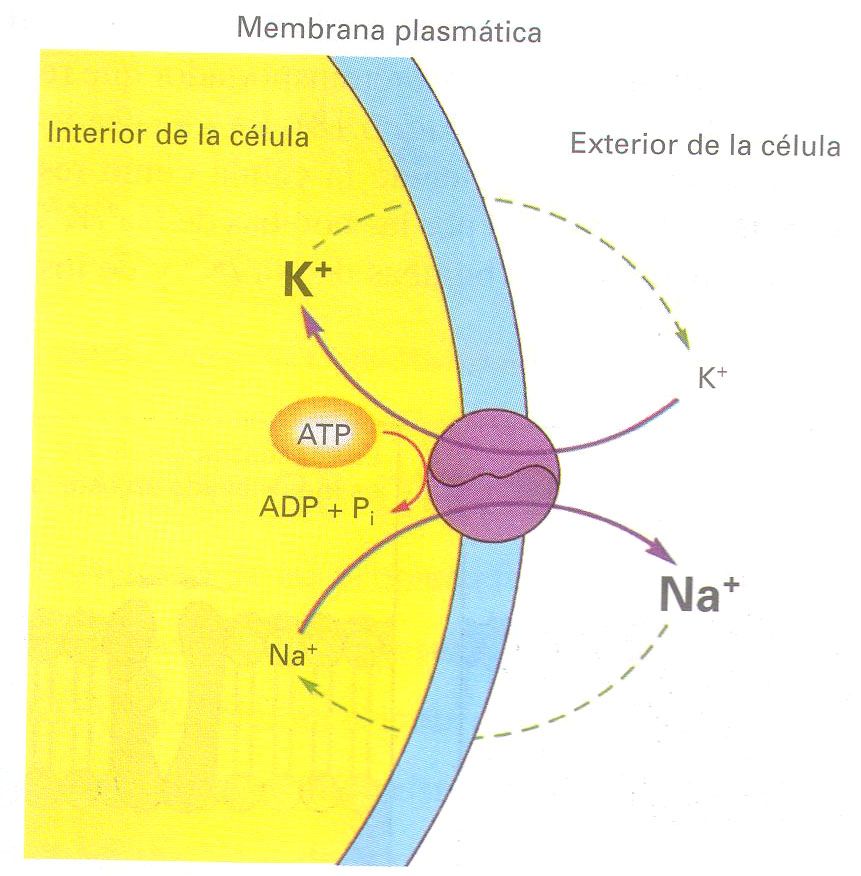
Potencial de membrana
Como resultado de las propiedades de permeabilidad de la membrana plasmática, la presencia de moléculas con carga negativa no difusibles en el interior de la células, y de la acción de las bombas de Na+/K+, existe una distribución desigual de cargas a ambos lados de la membrana. En consecuencia, el interior de la célula tiene carga negativa en comparación con el exterior. Esta diferencia de carga, o diferencia de potencial, se conoce como potencial de membrana.
El potencial de Nernst está definido como el nivel de potencial de difusión a través de una membrana que se opone directamente a la difusión neta de un ion en particular a través de la misma. Dicho potencial está en el interior de la membrana y se asume que el líquido extracelular se mantiene a un potencial eléctrico de cero voltios si la temperatura corporal es la adecuada (aproximadamente 37 °C).
Texto e imágenes: Fox, Stuart Ira. Fisiología humana séptima edición McGraw-Hill interamericana 2003, pp. 129-143
Los pigmentos fotosintéticos y la absorción de la luz
La clorofila está formada por un anillo porfirínico con un átomo de magnesio en el centro, asociado a un metanol y a un fitol (monoalcohol de compuesto de veinte carbonos). Como consecuencia, se conforma una molécula de carácter anfipático, en donde la porfirina actúa como polo hidrófilo y el fitol como polo lipófilo. Se distinguen dos variedades de clorofila: la clorofila a, que alberga un grupo metilo en el tercer carbono porfirínico y que absorbe luz de longitud de onda cercana a 630 nm, y la clorofila b, que contiene un grupo formilo y que absorbe a 660 nm.
Los carotenoides son isoprenoides y absorben luz de 440 nm, pudiendo ser de dos clases: los carotenos, que son de color rojo, y las xantófilas, derivados oxigenados de los nombrados anteriormente, que son de color amarillento. Las ficocianinas y las ficoeritrinas, de color azul y rojo respectivamente, son lípidos que se hayan asociados a proteínas originando las ficobiliproteínas.
Como los pigmentos fotosintéticos tienen enlaces covalentes sencillos que se alternan con enlaces covalentes dobles, se favorece la existencia de electrones libres que no pueden atribuirse a un átomo concreto.
Cuando incide un fotón sobre un electrón de un pigmento fotosintético de antena, el electrón capta la energía del fotón y asciende a posiciones más alejadas del núcleo atómico. En el supuesto caso de que el pigmento estuviese aislado, al descender al nivel inicial, la energía captada se liberaría en forma de calor o de radiación de mayor longitud de onda (fluorescencia). Sin embargo, al existir diversos tipos de pigmentos muy próximos, la energía de excitación captada por un determinado pigmento puede ser transferida a otro al que se induce el estado de excitación. Este fenómeno se produce gracias a un estado de resonancia entre la molécula dadora relajada y la aceptora. Para ello se necesita que el espectro de emisión del primero coincida, al menos en parte, con el de absorción del segundo. Los excitones se transfieren siempre hacia los pigmentos que absorben a mayor longitud de onda, continuando el proceso hasta alcanzar el pigmento fotosintético diana.
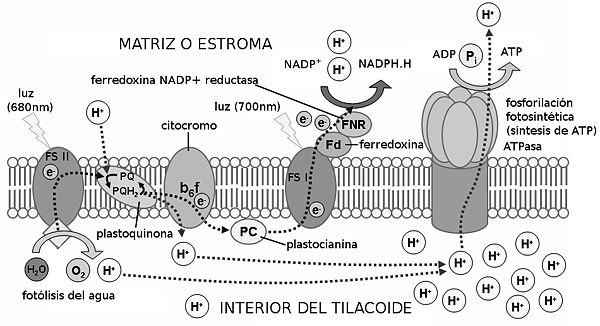
Fotofosforilación acíclica
Este proceso permite la formación de ATP y la reducción de NADP+ a NADPH + H+, y necesita de la energía de la luz, como ya se ha dicho. Se realiza gracias a los llamados fotosistemas, que se encuentran en la membrana de los tilacoides (en los cloroplastos). Estos están formados por dos partes:
• Antena, donde se agrupan los pigmentos antena, junto con proteínas, y cuya función es captar la energía de los fotones para transmitirla al pigmento diana; y el centro de reacción. Este está formado por proteínas y por pigmentos, encontrándose en él el llamado pigmento diana, que es aquel que recibe la energía de excitación de la antena, energía que sirve para excitar y liberar electrones. Aquí también se encuentra el primer dador de electrones, que repone los electrones al pigmento diana,
• Primer aceptor, que recibe los electrones liberados.
Hay dos tipos de fotosistemas:
• Fotosistema I, que se encuentra sobre todo en los tilacoides de estroma, y cuyo pigmento diana es la clorofila P700.
• Fotosistema II, que se encuentra sobre todo en los grana y cuyo pigmento diana es la clorofila P680.
Proceso
El proceso de la fase luminosa, supuesto para dos electrones, es el siguiente: Los fotones inciden sobre el fotosistema II, excitando y liberando dos electrones, que pasan al primer aceptor de electrones, la feofitina. Los electrones los repone el primer dador de electrones, el dador Z, con los electrones procedentes de la fotólisis del agua en el interior del tilacoide (la molécula de agua se divide en 2H+ + 2e- + 1/2O2). Los protones de la fotólisis se acumulan en el interior del tilacoide, y el oxígeno es liberado.
Los electrones pasan a una cadena de transporte de electrones, que invertirá su energía liberada en la síntesis de ATP. ¿Cómo? La teoría quimioosmótica nos lo explica de la siguiente manera: los electrones son cedidos a las plastoquinonas, las cuales captan también dos protones del estroma. Los electrones y los protones pasan al complejo de citocromos bf, que bombea los protones al interior del tilacoide. Se consigue así una gran concentración de protones en el tilacoide (entre éstos y los resultantes de la fotólisis del agua), que se compensa regresando al estroma a través de las proteínas ATP-sintasas, que invierten la energía del paso de los protones en sintetizar ATP. La síntesis de ATP en la fase fotoquímica se denomina fotofosforilación.
Los electrones de los citocromos pasan a la plastocianina, que los cede a su vez al fotosistema I. Con la energía de la luz, los electrones son de nuevo liberados y captados por el aceptor A0. De ahí pasan a través de una serie de filoquinonas hasta llegar a la ferredoxina. Ésta molécula los cede a la enzima NADP+-reductasa, que capta también dos protones del estroma. Con los dos protones y los dos electrones, reduce un NADP+ en NADPH + H+.
El balance final es: por cada molécula de agua (y por cada cuatro fotones) se forman media molécula de oxígeno, 1,3 moléculas de ATP, y un NADPH + H+.
Fase luminosa cíclica
En la fase luminosa o fotoquímica cíclica interviene de forma exclusiva el fotosistema I, generándose un flujo o ciclo de electrones que en cada vuelta da lugar a síntesis de ATP. Al no intervenir el fotosistema II, no hay fotólisis del agua y, por ende, no se produce la reducción del NADP+ ni se desprende oxígeno. Únicamente se obtiene ATP.
El objetivo que tiene la fase cíclica tratada es el de subsanar el déficit de ATP obtenido en la fase acíclica para poder afrontar la fase oscura posterior.
Cuando se ilumina con luz de longitud de onda superior a 680 nm (lo que se llama rojo lejano) sólo se produce el proceso cíclico. Al incidir los fotones sobre el fotosistema I, la clorofila P700 libera los electrones que llegan a la ferredoxina, la cual los cede a un citocromo b6 y éste a la plastoquinona (PH), que capta dos protones y pasa a (PQH2). La plastoquinona reducida cede los dos electrones al citocromo f e introduce
Tiene lugar al mismo tiempo que la acíclica. En ella sólo interviene el fotosistema I. Los electrones liberados, después de llegar a la ferredoxina, pasan a las plastoquinonas, y siguen la cadena de transporte de electrones hasta regresar a la plastocianina y al fotosistema I. Por tanto, se genera ATP pero no NADPH. Sirve para compensar el hecho de que en la fotofosforilación acíclica no se genera suficiente ATP para la fase oscura.
Analisis comparativo y evolutivo
La endosimbiosis es una asociación estrecha entre especies, en la que los individuos de una residen dentro de las células de la otra. Algunos orgánulos de las células eucariotas (células con núcleo), como las mitocondrias y los plastos (cloroplastos), proceden de su simbiosis inicial con ciertas bacterias.
Etimológicamente el término podría usarse para designar a cualquier simbionte que residiera en el interior del cuerpo de otro ser vivo, pero para este último concepto debe usarse el término endosomático (p.ej. simbionte endosomático). Éste es el caso, por ejemplo, de muchas de las bacterias que forman parte de la microbiota intestinal.
Los orgánulos de origen endosimbiótico aparecen muy transformados, pero conservan un genoma propio y se multiplican autónomamente, revelando su origen como organismos distintos.
Gracias a la endosimbiosis los organismos eucarióticos disfrutan de la capacidad de realizar procesos metabólicos que evolucionaron originalmente en bacterias. Es el caso de la respiración, de la que se ocupan las mitocondrias, la fotosíntesis, a cargo de los plastos o la fijación biológica de nitrógeno, realizada por bacterias, a menudo intracelulares, en las raíces de ciertas plantas.
En 1971 Lynn Margulis propuso la teoría de la endosimbiosis en serie, que explica la aparición de la célula eucariótica por asimilación simbiótica de varias bacterias con habilidades diferenciadas.
http://es.wikipedia.org/wiki/Endosimbiosis
http://www.textoscientificos.com/fotografia/fotoquimica
es.wikipedia.org/wiki/Fotosíntesis
Fotobiología
Fotoquímica
La fotoquímica es el estudio de las transformaciones químicas provocadas o catalizadas por la emisión o absorción de luz visible o radiación ultravioleta. Una molécula en su estado fundamental (no excitada) puede absorber un quantum de energía lumínica, esto produce una transición electrónica y la molécula pasa a un estado de mayor energía o estado excitado. Una molécula excitada es más reactiva que una molécula en su estado fundamental.
http://www.textoscientificos.com/fotografia/fotoquimica
Genoma mitocondrial y enfermedades genéticas del DNA mitocondrial
El genoma mitocondrial es una molécula de ADN circular que contiene 16.569 pares de bases y codifica 13 proteínas, 2 ARN ribosomal (ARNr) y 22 ARN de transferencia (ARNt). El código genético que utiliza es degenerado, es decir, ciertos codones en la mitocondria corresponden a aminoácidos diferentes de los utilizados por el genoma nuclear. Sin embargo, depende de muchas proteínas nucleares para poder replicarse y, a su vez, muchas proteínas presentes en las mitocondrias son codificadas por el genoma nuclear.
Enfermedades genéticas del DNA mitocondrial
Las enfermedades originadas por daños en el genoma mitocondrial tienen en común el estar producidas por una deficiencia en la biosíntesis de ATP, ya que toda la información que contiene este DNA está dirigida a la síntesis de proteínas componentes del sistema Oxphos.
Las manifestaciones de estas enfermedades son muy variadas y pueden afectar a todos los órganos y tejidos, ya que la síntesis de ATP se produce en todos ellos y a cualquier edad.
Estas pueden presentar una serie de aspectos clínicos, morfológicos y bioquímicos muy concretos que dan lugar a síndromes bien caracterizados pero, en la mayor parte de los casos, principalmente en edad pediátrica, los síntomas son muy poco informativos y es sólo la presencia de anormalidades neurológicas, a veces acompañadas de aumento de ácido láctico y de otros síntomas clínicos secundarios que afectan a diversos órganos, lo que da alguna orientación en el diagnóstico de una enfermedad mitocondrial. Entre las manifestaciones clínicas más comunes se encuentran una o varias de las siguientes: desórdenes motores, accidentes cerebrovasculares, convulsiones, demencia, intolerancia al ejercicio, oftalmoplejia, retinopatía pigmentaria, atrofia óptica, ceguera, sordera, cardiomiopatía, disfunciones hepáticas y pancreáticas, diabetes, defectos de crecimiento, anemia sideroblástica, pseudo obstrucción intestinal, nefropatías, acidosis metabólica y otras más secundarias.
http://es.wikipedia.org/wiki/Genoma_mitocondrial
http://www.insp.mx/rsp/_files/File/2001/VOL%2043%202/v43_2_enfermedades%20geneticas.pdf
Respiración Mitocondrial
ELECTRODO DE OXÍGENO
El electrodo de oxígeno comprende un cátodo de platino central (B) unido a una resina y un ánodo de plata (C) concéntrico unido por un puente electrolítico y conectados al módulo control.
La cámara del electrodo es preparada por aplicación de un espaciador de papel muy fino y una fina membrana de poli-tetra-fluor-etileno (P.T.F.E.) que es cuidadosamente fijada a la placa base donde se encuentran los electrodos por un anillo-O. En la presencia de oxígeno una pequeña corriente fluye a través de los electrodos que es proporcional a la concentración de oxígeno en la muestra. Esta señal es digitalizada por la unidad de control y presentada directamente en el PC.
Estos electrodos pueden ser acondicionados para medidas en fase líquida o en fase gaseosa; aquí vamos a medir en fase líquida.
Todas las unidades del electrodo deben mantenerse a temperatura constante durante las determinaciones. Este efecto se consigue por circulación de agua a la temperatura deseada alrededor de la cámara y controlando la temperatura de los componentes de la muestra. Este control es importante por dos razones:
1º.- El electrodo es sensible a la temperatura
2º.- El contenido en oxígeno de las muestras acuosas saturadas de aire cambia con la temperatura.
http://www2.uah.es/tejedor_bio/bioquimica_ambiental/guion-res-mitocondrial.pdf
Desacopladores e inhibidores.
Gran parte del conocimiento de la función mitocondrial ha resultado de estudios con compuestos tóxicos. Inhibidores específicos se han usado para distinguir el sistema de transporte de electrones del sistema de fosforilación oxidativa, y ha ayudado a definir la secuencia de los transportadores redox en la cadena. Si la cadena se bloquea en un punto, todos los transportadores anteriores quedan más reducidos, y los posteriores más oxidados.
La rotenona es una toxina de una planta, utilizada por indios amazónicos como veneno, también ha sido usada como insecticida.

El amital (barbitúrico) inhibe al complejo I, afecta las oxidaciones dependientes del NAD+.
La antimicina A (Antibiótico).
Actúa a inhibiendo el complejo III. Inhibe la reoxidación del NADH y del FADH2.
El cianuro bloquea el paso de electrones del citocromo a3 al oxígeno.
Estos inhibidores detienen el paso de electrones de modo que no hay bombeo de protones. Sin gradiente de protones, no hay síntesis de ATP.
La oligomicina, un antibiótico producido por Streptomyces, inhibe a la ATPasa al unirse a la subunidad Fo e interferir en el transporte de H+ a través de Fo, inhibe por lo tanto la síntesis de ATP.
Diciclohexilcarbodiimida (DCCD), un reactivo soluble en lípidos, también inhibe el transporte de protones por Fo al reaccionar con un residuo de glutámico en una de las subunidades de Fo de mamíferos.
En estas condiciones el gradiente de protones que se produce es mayor que lo normal, sin embargo la energía potencial de éste no puede ser utilizada para producir ATP.
Estos venenos, como el 2,4 dinitrofenol (DNP), el carbonilcianuro-p-trifluorometoxi-hidrazona (FCCP) y el carbonilcianuro-m-clorofenilhidrazona (CCCP) desacoplan la fosforilación oxidativa de la cadena respiratoria, se conocen como agentes desacopladores.

La forma protonada, sin carga eléctrica de estos compuestos, pasa a través de la membrana interna mitocondrial intacta, descargando así el gradiente de pH. En la matriz, a pH más bajo, el ácido débil se disocia, la forma disociada pasa la membrana interna, destruyendo el potencial de membrana. Este proceso se puede repetir, de modo que una pequeña cantidad del agente desacoplante puede catalizar el paso de una cantidad enorme de protones y hacer un corto circuito en la cadena respiratoria.
En resumen, permitiendo el paso de protones a través de la membrana, se disipa el gradiente de protones, no hay bombeo de protones a través de la ATP-sintasa con producción de ATP.
Los agentes desacoplantes son todos sintéticos, sin embargo en el mitocondria del tejido adiposo pardo una proteína desacopladora (termogenina) participa en el delicado control de la termogénesis.
4. Inhibidores de transporte (atractalósido) que previenen ya sea la salida del ATP o la entrada de material combustible a través de la membrana mitocondrial interna.

5. Ionósforos (valinomicina, nigericina) que permiten el paso a través de la membrana a compuestos que normalmente están impedidos.
6. Inhibidores del ciclo de Krebs (arsenito) que bloquean una o más enzimas del ciclo de Krebs.
La producción de ATP aeróbica es más eficiente que la producción anaeróbica.
Glicólisis anaeróbica:
C6H12O6 + 2ADP + 2Pi -> 2 lactato + 2H+ + 2ATP
C6H12O6 + 38ADP + 38Pi 6 O2 -> 6 CO2 + 44H20 + 38ATP
Por otra parte, el cálculo tradicional de 36 ATP o de 38 ATP, según funcione la lanzadera del glicerofosfato o del malato respectivamente, es obsoleto. Mediciones recientes estiman 30 ATP por glucosa totalmente metabolizada.
http://www.bioquimicaqui11601.ucv.cl/unidades/cte/traselectfofox4fid.html
Bioenergética mitocondrial
La teoría quimiosmótica enunciada por Peter Mitchell, explica cómo la energía derivada del transporte de electrones por la cadena de transporte de electrones se utiliza para producir ATP a partir de ADP y Pi.
La bomba de protones: el transporte de electrones está acoplado al transporte de H+ a través de la membrana interna mitocondrial desde el espacio intermembranal.
Este proceso crea simultáneamente a través de la membrana interna mitocondrial un gradiente eléctrico (con más cargas positivas en el exterior de la membrana que en la matriz mitocondrial) y un gradiente de pH (el exterior de la membrana está a un pH más ácido que el interior). La energía generada por este gradiente es suficiente para realizar la síntesis de ATP.
La transferencia de electrones en la cadena de transporte de electrones es energéticamente favorable porque el NADH es un poderoso donador de electrones y el Oxígeno molecular es un potente aceptor de electrones. De hecho el flujo neto de electrones desde el NADH hasta el Oxígeno resulta en la síntesis de ATP. La fosforilación oxidativa es una serie de eventos químicos que llevan a la síntesis de ATP:
ADP + Pi síntesis del ATP
fosforilación del ADP
El evento vital se lleva a cabo en la membrana plasmática bacteriana, en la membrana interna mitocondrial y en los tilacoides de los cloroplastos.
En la década de los 30´s: Belitzer y Tsivakoba encontraron que el proceso de la fosforilación de ADP en los tejidos animales estaba asociado a la respiración o consumo de O2. Más adelante se describió que la respiración se lleva a cabo en las mitocondrias.
Krebs encontró el ciclo de los ácidos tricarboxílicos en el cual el piruvato se transforma en Ac-CoA que a su vez interviene en la reducción de NAD+ y en la posterior generación del succinato.
Como ha sucedido muchas veces a los largo de la historia de la investigación científica, dos investigadores reportaron simultáneamente un evento bioquímico. En 1937, Kalkar en Dinamarca y Belitzer en la antigua URSS, encontraron una correlación muy interesante entre la desaparición del Pi y la respiración. Estudiaron el efecto de la adición de Pi (HPO34) a homogenados de tejidos de mamíferos; el experimento lo realizaron en presencia y ausencia de 02 o en presencia de cianuro (CN¯). Reportaron que a medida que se consumía el 02 el Pi desaparecía del medio de reacción y que cuando agregaban a un inhibidor del consumo de 02, CN¯ e este caso, el proceso no se llevaba a cabo. Posteriormente se verificó que la síntesis de ATP es una reacción endergonica, en la cual la respiración o consumo de 02 acopladas a la fosforilación del ADP, genera energía.
En los seres vivos la oxidación de moléculas orgánicas tiene como resultado el movimiento de protones (H+) del interior de la matriz mitocondrial al espacio intermembranal en mitocondrias y cloroplastos o bien al citoplasma en las bacterias. La cadena de transporte de electrones y la fosforilación oxidativa estuvieron separadas conceptualmente por mucho tiempo. Las observaciones de la formación del ATP hacían pensar a los investigadores en buscaba un intermediario fosforilado de la reación. Hasta que en 1961 Peter Mitchell propuso la hipótesis quimiosmótica en la cual propuso que el intermediario energético necesario para la formación del ATP (o fosforilación del ADP), era una diferencia en la concentración de protones a través de la membrana.
Gracias a estas observaciones Mitchell recibió en premio Nobel de Química en 1978. Murió al final de la década de los 80´s.
http://laguna.fmedic.unam.mx/~evazquez/0403/fosforilacion%20oxidativa.html
Reacciones oxido-reducción
Potencial Redox
También llamado potencial de reducción es un concepto electroquímico que se puede determinar midiendo la actividad de los electrones.
Radicales libres
Los radicales libres son resultado de los procesos fisiológicos propios del organismo, como el metabolismo de los alimentos, la respiración y el ejercicio, o bien son generados por factores ambientales como la contaminación industrial, el tabaco, la radiación, los medicamentos, los aditivos químicos en alimentos procesados y los pesticidas. Son átomos o moléculas extremadamente reactivas, debido a que en el orbital más externo de su estructura tienen uno o más electrones sin aparear. Esta inestabilidad les confiere una avidez física por la captura de un electrón de cualquier otra molécula de su entorno, ocasionando que la estructura afectada quede inestable. De esta forma pueden establecer reacciones en cadena por medio de varios transportadores que se oxidan y se reducen secuencialmente, cuando un radical libre inicial modifica una biomolécula después de transferir o capturar un electrón. El daño es transmitido por medio de los transportadores, que incluso pueden ser moléculas circulantes.

Berg, Jeremy M. Tymoczko, John L. Stryer, Lumbert. Bioquímica 6a edición, España, Reverté, 2008 pag. 506
imagen: http://www.alumno.unam.mx/algo_leer/Envejecimiento.pdf
ATP

El ATP es la divisa o unidad biológica universal de energía. El elevado potencial para transferir fosforilos del ATP capacita para utilizarse como fuente de energía en la contracción muscular, el trasporte activo, la amplificación de señales y la biosíntesis. En la célula, la hidrólisis de la molécula de ATP cambia la relación de equilibrio entre productos y sustancias reaccionantes de una reacción acoplada. De esta forma, una secuencia de reacciones termodinámicamente desfavorable puede resultar muy favorable si se acopla a la hidrólisis de un número suficiente de moléculas de ATP.
El ATP se genera en la combustión de moléculas combustibles como la glucosa, los ácidos grasos y aminoácidos. El intermediario común en la mayoría de estas oxidaciones es el acetil-CoA. Los átomos del carbono del fragmento acetilo se oxidan por completo a CO2 por el ciclo de Krebs con formación simultánea de NADH y FADH2. Estos transportadores de electrones ceden sus electrones de elevado potencial a la cadena respiratoria. El subsiguiente flujo de electrones al O2 origina el bombeo de protones a través de la membrana interna mitocondrial. El gradiente de protones se emplea con posterioridad para sintetizar ATP. La glucolisis también genera ATP, pero la cantidad formada es mucho menor que la producida por la fosforilación oxidativa. La oxidación de glucosa a piruvato genera 2 moléculas de ATP, mientras que si la glucosa se oxida por completo a CO2 se generan 30 moléculas de ATP.
El NADH es el principal dador de electrones en las biosíntesis reductora. En la mayoría de las biosíntesis los productos están más reducidos que sus precursores y por eso se necesita, además de ATP, poder reductor. Los electrones de elevado potencial que se requieren para dirigir estas reacciones proceden normalmente del NADPH.
Las biomoléculas se construyen apartir de una serie pequeña de precursores. La enorme variedad de moléculas de los seres vivos se sintetizan a partir de un número mucho menor de precursores. Las vías metabolicas, que generan ATP y NADH, también proporcionan precursores para la biosíntesis de moléculas complejas.
Las vías biosinteticas y degradativas casi siempre son diferentes. Asi, por ejemplo, la vía de síntesis de los ácidos grasos es diferente de la de su degradación. Esta separación posibilita que las vías biosinteticas y degradativas sean termodinámicamente favorables en todo momento. Una vía biosintetica se convierte en exergonica si se acopla a la hidrólisis de un número suficente de moléculas de ATP.
En la hidrólisis del ATP se está hidrolizando uno de los enlaces anhídrido de ácido. Esto libera gran energía, concretamente ΔG = -7,7 kcal/mol
Las razones químicas de esa tendencia son tres:
En la célula existen muchos enlaces de alta energía, la mayoría de los cuales son enlaces fosfato. El ATP ocupa una posición intermedia entre los fosfatos de alta energía.
{kind=link}
Trabajo Biológico

Giancoli, Douglas C. Física principios con aplicación 6ª edición, Pearson educación, 2006 pp. 414
Fotografía: http://www.bonkos.com/pix/2006/04/gatorade2.jpg
Transducción de energía
Otras veces, este potencial es la respuesta al estímulo que se da en una región determinada de una célula, como es el caso del Corpúsculo de Pacini. Este es un terminal nervioso encapsulado que en respuesta a un estímulo mecánico genera un potencial local, graduado, que induce en el primer nódulo de Ranvier de ese axón, potenciales de acción propagados. A estos potenciales locales capaces de producir potenciales de acción se les llama también potenciales generadores.
http://www.uc.cl/sw_educ/neurociencias/html/111.html
Energía libre de Gibbs
La segunda ley de la termodinámica postula que una reacción química espontánea hace que la entropía del universo aumente, ΔS universo > 0, así mismo ΔS universo está en función de ΔS sistema y ΔS alrededores. Por lo general sólo importa lo que ocurre en el sistema en estudio y; por otro lado el cálculo de ΔS alrededores puede ser complicado.
Por esta razón fue necesario otra función termodinámica, la energía libre de Gibbs, que sirva para calcular si una reacción ocurre de forma espontánea tomando en cuenta solo las variables del sistema.
Cálculo de la energía libre de Gibbs
Los cambios en la energía libre
Contenido de calor; T es la temperatura y S es la entropía del sistema. Fue desarrollada en los años 1870 por el físico-matemático estadounidense Williard Gibbs.
Cambios de energía libre estándar
La energía libre de reacción, se denota, ΔG reacción, es el cambio de energía en una reacción a condiciones estandares. En esta reacción los reactivos en su estado estandar se convierten en productos en su estado estandar.
Dada la siguiente ecuación química:
aA+bB -> cC+dD
La energía libre se calcula como

Donde A y B son los reactivos en estado estandar y; C y D son los productos en su estado estandar. Además a, b, c y d son sus respectivos coeficientes estequiométricos.
en general:

donde m y n son los coeficientes estequiométricos.
Así como en el cálculo de la entalpía, en la energía libre estándar de formación para cualquier elemento en su forma estable (1 atm y 25ºC) es 0
La variación de energía libre de Gibbs para un proceso a temperatura constante viene dada por:

La temperatura puede ser un factor determinante a la hora de hacer que un proceso sea espontaneo o no lo sea.
es.wikipedia.org/wiki/Energía_libre_de_Gibbs
Constante de equilibrio
Reactivos <-> Productos Kc=([productos]/
([reactivos])
Garritz, A. Chamizo, J.A. Química México Pearson educación, 1998 pp. 639-640
Nociones de Termodinámica
El calor es muy parecido al trabajo, para distiguirlos el calor se define como una transferencia de energía provocada por una diferencia de temperatura, mientras que el trabajo es una transferencia de energía que no se debe a una diferencia de temperatura.
En la termodinámica, con frecuencia se hace referencia a sistemas particulares. Un sistema es cualquier objeto o conjunto de objetos que se desean considerar, al resto del universo se le considera como el ambiente o los alrededores.
La primera ley de la termodinámica relaciona el trabajo y la transferencia de calor con el intercambio de energía interna de un sistema, y se refiere a la conservación de la energía.
La segunda ley de la termodinámica expresa límites en la capacidad para realizar trabajo útil y con frecuencia se establece en términos de entropía.
Giancoli, Douglas C. Física principios con aplicación 6ª edición, Pearson educación, 2006 pp. 408-416
Introducción a la Bioenergética
La bioenergética es la parte de la biología que está relacionada con la física y que se encarga del estudio de los procesos de absorción, transformación y entrega de energía en los sistemas biológicos.
En general, la bioenergética se relaciona con la termodinámica, en particular con la energía libre y la energía libre de Gibbs. Los cambios en la energía libre de Gibbs ΔG nos dan una cuantificación de la factibilidad energética de una reacción química y pueden proveer de una predicción de si la reacción podrá suceder o no. Como una característica general de la bioenergética, esta solo se interesa por los estados energéticos inicial y final de los componentes de una reacción química, los tiempos necesarios para que el cambio químico se lleve a cabo en general se desprecian. Un objetivo general de la Bioenergética es predecir si ciertos procesos son posibles o no; en general la cinética cuantifica qué tan rápido ocurre la reacción química.