La clorofila está formada por un anillo porfirínico con un átomo de magnesio en el centro, asociado a un metanol y a un fitol (monoalcohol de compuesto de veinte carbonos). Como consecuencia, se conforma una molécula de carácter anfipático, en donde la porfirina actúa como polo hidrófilo y el fitol como polo lipófilo. Se distinguen dos variedades de clorofila: la clorofila a, que alberga un grupo metilo en el tercer carbono porfirínico y que absorbe luz de longitud de onda cercana a 630 nm, y la clorofila b, que contiene un grupo formilo y que absorbe a 660 nm.
Los carotenoides son isoprenoides y absorben luz de 440 nm, pudiendo ser de dos clases: los carotenos, que son de color rojo, y las xantófilas, derivados oxigenados de los nombrados anteriormente, que son de color amarillento. Las ficocianinas y las ficoeritrinas, de color azul y rojo respectivamente, son lípidos que se hayan asociados a proteínas originando las ficobiliproteínas.
Como los pigmentos fotosintéticos tienen enlaces covalentes sencillos que se alternan con enlaces covalentes dobles, se favorece la existencia de electrones libres que no pueden atribuirse a un átomo concreto.
Cuando incide un fotón sobre un electrón de un pigmento fotosintético de antena, el electrón capta la energía del fotón y asciende a posiciones más alejadas del núcleo atómico. En el supuesto caso de que el pigmento estuviese aislado, al descender al nivel inicial, la energía captada se liberaría en forma de calor o de radiación de mayor longitud de onda (fluorescencia). Sin embargo, al existir diversos tipos de pigmentos muy próximos, la energía de excitación captada por un determinado pigmento puede ser transferida a otro al que se induce el estado de excitación. Este fenómeno se produce gracias a un estado de resonancia entre la molécula dadora relajada y la aceptora. Para ello se necesita que el espectro de emisión del primero coincida, al menos en parte, con el de absorción del segundo. Los excitones se transfieren siempre hacia los pigmentos que absorben a mayor longitud de onda, continuando el proceso hasta alcanzar el pigmento fotosintético diana.
Fotofosforilación acíclica
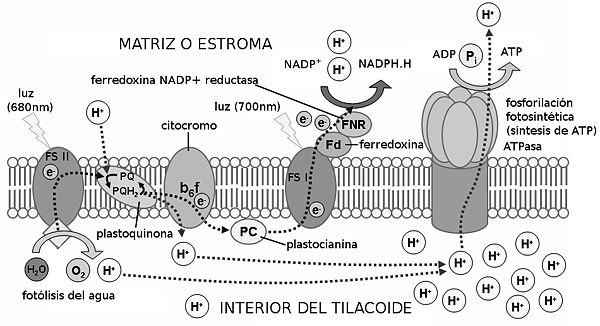
Este proceso permite la formación de ATP y la reducción de NADP+ a NADPH + H+, y necesita de la energía de la luz, como ya se ha dicho. Se realiza gracias a los llamados fotosistemas, que se encuentran en la membrana de los tilacoides (en los cloroplastos). Estos están formados por dos partes:
• Antena, donde se agrupan los pigmentos antena, junto con proteínas, y cuya función es captar la energía de los fotones para transmitirla al pigmento diana; y el centro de reacción. Este está formado por proteínas y por pigmentos, encontrándose en él el llamado pigmento diana, que es aquel que recibe la energía de excitación de la antena, energía que sirve para excitar y liberar electrones. Aquí también se encuentra el primer dador de electrones, que repone los electrones al pigmento diana,
• Primer aceptor, que recibe los electrones liberados.
Hay dos tipos de fotosistemas:
• Fotosistema I, que se encuentra sobre todo en los tilacoides de estroma, y cuyo pigmento diana es la clorofila P700.
• Fotosistema II, que se encuentra sobre todo en los grana y cuyo pigmento diana es la clorofila P680.
Proceso
El proceso de la fase luminosa, supuesto para dos electrones, es el siguiente: Los fotones inciden sobre el fotosistema II, excitando y liberando dos electrones, que pasan al primer aceptor de electrones, la feofitina. Los electrones los repone el primer dador de electrones, el dador Z, con los electrones procedentes de la fotólisis del agua en el interior del tilacoide (la molécula de agua se divide en 2H+ + 2e- + 1/2O2). Los protones de la fotólisis se acumulan en el interior del tilacoide, y el oxígeno es liberado.
Los electrones pasan a una cadena de transporte de electrones, que invertirá su energía liberada en la síntesis de ATP. ¿Cómo? La teoría quimioosmótica nos lo explica de la siguiente manera: los electrones son cedidos a las plastoquinonas, las cuales captan también dos protones del estroma. Los electrones y los protones pasan al complejo de citocromos bf, que bombea los protones al interior del tilacoide. Se consigue así una gran concentración de protones en el tilacoide (entre éstos y los resultantes de la fotólisis del agua), que se compensa regresando al estroma a través de las proteínas ATP-sintasas, que invierten la energía del paso de los protones en sintetizar ATP. La síntesis de ATP en la fase fotoquímica se denomina fotofosforilación.
Los electrones de los citocromos pasan a la plastocianina, que los cede a su vez al fotosistema I. Con la energía de la luz, los electrones son de nuevo liberados y captados por el aceptor A0. De ahí pasan a través de una serie de filoquinonas hasta llegar a la ferredoxina. Ésta molécula los cede a la enzima NADP+-reductasa, que capta también dos protones del estroma. Con los dos protones y los dos electrones, reduce un NADP+ en NADPH + H+.
El balance final es: por cada molécula de agua (y por cada cuatro fotones) se forman media molécula de oxígeno, 1,3 moléculas de ATP, y un NADPH + H+.
Fase luminosa cíclica
En la fase luminosa o fotoquímica cíclica interviene de forma exclusiva el fotosistema I, generándose un flujo o ciclo de electrones que en cada vuelta da lugar a síntesis de ATP. Al no intervenir el fotosistema II, no hay fotólisis del agua y, por ende, no se produce la reducción del NADP+ ni se desprende oxígeno. Únicamente se obtiene ATP.
El objetivo que tiene la fase cíclica tratada es el de subsanar el déficit de ATP obtenido en la fase acíclica para poder afrontar la fase oscura posterior.
Cuando se ilumina con luz de longitud de onda superior a 680 nm (lo que se llama rojo lejano) sólo se produce el proceso cíclico. Al incidir los fotones sobre el fotosistema I, la clorofila P700 libera los electrones que llegan a la ferredoxina, la cual los cede a un citocromo b6 y éste a la plastoquinona (PH), que capta dos protones y pasa a (PQH2). La plastoquinona reducida cede los dos electrones al citocromo f e introduce
Tiene lugar al mismo tiempo que la acíclica. En ella sólo interviene el fotosistema I. Los electrones liberados, después de llegar a la ferredoxina, pasan a las plastoquinonas, y siguen la cadena de transporte de electrones hasta regresar a la plastocianina y al fotosistema I. Por tanto, se genera ATP pero no NADPH. Sirve para compensar el hecho de que en la fotofosforilación acíclica no se genera suficiente ATP para la fase oscura.
Analisis comparativo y evolutivo
La endosimbiosis es una asociación estrecha entre especies, en la que los individuos de una residen dentro de las células de la otra. Algunos orgánulos de las células eucariotas (células con núcleo), como las mitocondrias y los plastos (cloroplastos), proceden de su simbiosis inicial con ciertas bacterias.
Etimológicamente el término podría usarse para designar a cualquier simbionte que residiera en el interior del cuerpo de otro ser vivo, pero para este último concepto debe usarse el término endosomático (p.ej. simbionte endosomático). Éste es el caso, por ejemplo, de muchas de las bacterias que forman parte de la microbiota intestinal.
Los orgánulos de origen endosimbiótico aparecen muy transformados, pero conservan un genoma propio y se multiplican autónomamente, revelando su origen como organismos distintos.
Gracias a la endosimbiosis los organismos eucarióticos disfrutan de la capacidad de realizar procesos metabólicos que evolucionaron originalmente en bacterias. Es el caso de la respiración, de la que se ocupan las mitocondrias, la fotosíntesis, a cargo de los plastos o la fijación biológica de nitrógeno, realizada por bacterias, a menudo intracelulares, en las raíces de ciertas plantas.
En 1971 Lynn Margulis propuso la teoría de la endosimbiosis en serie, que explica la aparición de la célula eucariótica por asimilación simbiótica de varias bacterias con habilidades diferenciadas.
http://es.wikipedia.org/wiki/Endosimbiosis
http://www.textoscientificos.com/fotografia/fotoquimica
es.wikipedia.org/wiki/Fotosíntesis
No hay comentarios:
Publicar un comentario